


1 前言
嫩江流域是指嫩江及其支流流经的地域,包括内蒙古自治区呼伦贝尔盟、兴安盟、黑龙江省齐齐哈尔市、大庆以及吉林省白城地区。它的西面和北面被高耸的大兴安岭与小兴安岭所围绕,东面是丘陵起伏的张广才岭,南接松辽大平原,处于三面环山的低地。嫩江流域地貌复杂多样,包括山地、丘陵、平原、沼泽、湿地等,水资源非常丰富,自古以来就是我国东北古代先民的重要栖息地之一。作为一个相对封闭的地理区域,嫩江流域形成了其独特而又连续的考古文化序列,从新石器时代早期的后套木嘎一期文化、双塔一期文化、到新石器时代中期的黄家围子文化、后套木嘎三期文化,再到新石器时代晚期的哈民忙哈文化、昂昂溪文化,整个地区的新石器文化面貌显示出较大的一致性和继承性,以大量的压制细石器和渔猎工具为特征,同时存在大量野生动物骨骼、房址、环壕等,展现出非常鲜明的定居式的渔猎经济文化类型[1,2]。大约在距今4000年,伴随着新石器时代晚期的昂昂溪文化在嫩江中游的消失,嫩江流域的新石器时代文化结束,随之兴起的是嫩江中下游的青铜铁器文化[3]。尽管嫩江流域的青铜文化与新石器文化有不同程度的递进和演化关系,渔猎经济在先民生活中仍然占用重要地位,但是,马、牛、羊、猪和狗等家畜骨骼的存在、丰富的禾本科植物遗存、大量的磨盘和磨棒,表明这一时期的先民开始兼营畜牧业和少量的种植业[4,5,6]。
根据古气候学、古环境学研究,嫩江流域的气候环境在距今5000-3000年期间,处于一个气温持续降低的过程,气候明显恶化,草本植物比例增加,木本植物比例减少,影响了人们赖以生存的动植物资源分布[3]。嫩江流域青铜早期畜牧业和种植业的出现可能就是人们为应对这一气候变化所做出的调整与改变。此外,这一时期,嫩江流域的先民与外界的联系不断加强。考古学研究表明,吉林后套木嘎四期出现的玉器种类和形式以及部分陶器的纹饰均受到了辽西地区红山文化的影响,而陶器的主体特征则受到呼伦贝尔草原的影响较为明显[6],昂昂溪五福墓地出土的陶罐与骨鱼镖则与贝加尔湖地区的相似[5];语言学证据支持,新石器晚期至青铜时期盛行于嫩江流域的泛欧亚语言来源于西辽河地区[7]。嫩江流域青铜时期以来,与外来文化的联系是其自身为了适应环境的改变向周边古代人群引进了先进的文化技术,还是周边古代人群迫于生存压力向嫩江流域迁徙带去了先进的文化技术,一直是考古学关注的焦点。
应用遗传学方法从嫩江流域古代人类遗骸中获取其遗传信息,通过对比新石器时代与青铜铁器时代人群的遗传结构变化与周边对应时期人群的遗传信息,是解决以上问题的一种有效方法。我们对已发表的嫩江流域吉林后套木嘎遗址[8]、黑龙江洪河村遗址[7]、内蒙古扎赉诺尔[9]遗址等10个个体的全基因组数据进行分析发现,嫩江流域早晚期人群在全基因上展现了较强的遗传连续性,只有一个晚期个体携带少量的蒙古/北亚成分,并没有发现其他地区古代人群的遗传成分的进入。众所周知,常染色体会在减数分裂期发生重组,造成基因组等位基因频率的改变,当外来的基因成分比例较低,经过多代繁衍以后,某些基因突变会被稀释到难以检测的水平,因此可能会掩盖部分外来的基因成分。此外,常染色体突变率低,当两个遗传结构相似的种群发生基因交流时很难被检测到。而作为单亲遗传标记,线粒体DNA因为不发生重组,有利于部分突变快速的分离并得到富集,使得某些变异在线粒体基因组水平更容易检测到。此外,线粒体DNA突变率高,种群内部差异显著,具有更明显的地域性分布特性,在追踪人群来源、评估人群扩张历史方面具有其独特的优势[10]。
2 材料和方法
2.1. 材料
本研究对松嫩平原新石器至铁器时代的6个考古遗址的24个样本进行了收集。新石器时代个体收集自四个遗址,分别是吉林大安后套木嘎遗址(6个)、内蒙古五七遗址(1个)、黑龙江洪河村遗址(4个)以及内蒙古扎赉诺尔遗址新石器时代文化层(1个);青铜及早期铁器时代的个体收集于吉林大安后套木嘎汉书二期(6个)、内蒙古蘑菇山遗址(3个)、黑龙江大古堆遗址(2个)、内蒙古扎赉诺尔遗址铁器时代文化层(1个)四个遗址。其中,五七农场遗址、洪河村遗址、扎赉诺尔遗址、蘑菇山遗址的个体线粒体基因组是从已发表得全基因组shotgun数据中分离产生[7,9]。其他个体的线粒体基因组使用线粒体捕获探针富集后测序产生。根据碳十四测序数据,本次研究的24个样本年代跨度约1万年,从新石器时代的11000 BP到铁器时代2000 BP,新石器时代个体(11000-4000BP)数量为12个,青铜及早期铁器时代个体(4000BP-2000 BP)数量为12个。详细的遗址位置信息见图1,样本详细信息见表1。
图1
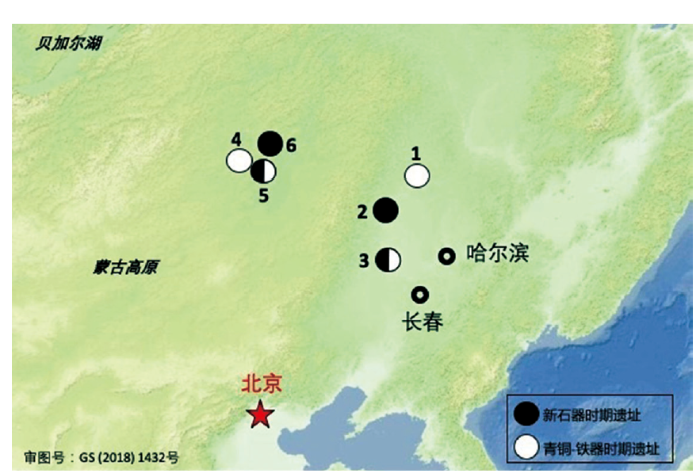
图1
本研究所收集古代样本的遗址地理位置
1.大古堆遗址;2.洪河村遗址;3.后套木嘎遗址;4.蘑菇山鲜卑遗址;5.扎赉诺尔遗址;6.五七农场遗址
Fig.1
Geographic map showing the sampling locations.
表1 本研究中考古样本背景信息
Tab.1
| 遗址名称 | 年代(cal BP) | 样本数量(个) | 年代参考文献 | |
|---|---|---|---|---|
| 新石器时代 | 后套木嘎遗址 | 11000-6000 | 6 | [8] |
| 扎赉诺尔遗址 | 5500-5300 | 1 | [9] | |
| 黑龙江五七农场遗址 | 5500-5300 | 1 | [9] | |
| 洪河村遗址 | 4000 | 4 | [7] | |
| 青铜铁器时代 | 后套木嘎遗址 | 4000-2000 | 6 | [8] |
| 黑龙江大古堆遗址 | 2500-2000 | 2 | [16] | |
| 内蒙古蘑菇山遗址 | 2000 | 3 | [9] | |
| 扎赉诺尔遗址 | 2000-2200 | 1 | [9] |
2.2. 实验方法
1) 古DNA的提取、建库和测序 在专业的考古DNA实验室中进行了样本制备、骨骼和牙齿样本的 DNA 提取以及文库制备。 首先应用DNA-off和10%次氯酸钠溶液擦拭和浸泡牙齿5分钟,然后在超净工作台中应用254nm的紫外线照射牙齿表面各30分钟,钻取牙粉50毫克至500毫克不等。 应用EDTA(0.5 M)和蛋白酶 K (10mg / mL) 50°C消化过夜,使用minElut 纯化试剂盒进行纯化,最后的洗脱体系为60微升。在DNA提取中,每7个样品加入一个空白阴性对照。经过线粒体高可变区部分片段PCR扩增评估样本提取效率和洁净度。应用宁超等人描述的方法进行双链DNA建库[8]。使用商业公司的illumina Hiseq x10 平台进行序列分析。
2) 序列评估和真实性验证 参考宁超等人的方法进行序列分析[8]。首先去掉接头和末端损伤,将DNA片段映射到人类参考基因组(版本 hs37d5)上,并过滤掉10%不匹配的以及片段长度小于30bp 的reads,通过损伤检验来评估古DNA的错配模式,通过calico.0.2. py(
3) 统计学分析 我们首先将去掉接头序列的FASTQ文件与人类基因组参考序列hs37d5.fa进行比对,并利用SAMtools去除低质量的序列(-q30)。接下来我们用mapDamage软件进行古DNA末端损伤鉴定,再利用bamUtils软件中的trimBam命令将序列两端具有损伤模式的8bp碱基切掉。最后,我们利用SAMtools软件里view命令将比对到线粒体上的序列进行提取,得到BAM文件。接下来我们使用Geneious软件(
为了分析西辽河和嫩江流域地区的人群历史和有效人口大小,我们利用BEAST2软件进行Extended Bayesian Skyline Plot(EBSP)分析。分析的对象主要包括N9a和B4c1两个单倍群,其中N9a属于两个地区的共享谱系,而B4c1在嫩江流域新石器和青铜时期人群中都有发现。其中N9a的分析包括71个现代个体,B4c1类型的分析包括44个现代个体。EBSP分析采用HKY碱基替代模型和严格分子钟方法,线粒体全序的分子钟速率设置为2.53×10-8(由4.5万年前Ust-Ishim推导出)[16]。将使用MEGA软件构建的最大简约树作为起始树导出谱系树,估计各个线粒体谱系分支的共祖时间,得到贝叶斯人口变化曲线。此外,为了进一步探讨谱系内个体之间的关系,我们应用Network软件对单倍群B4c1a2以及N9a执行了中介网络分析。
3 结果
我们共获得24个样本的线粒体全序列,数据覆盖率在4.7-853.4X之间,污染率(0.6-9.2%)均很低。这24个个体共包含24个单倍型,可以归属到10个线粒体单倍群:A、B、C、D、F、G、N9a、M8、Z、Y, 均属于东部欧亚谱系(表2)。为了追踪嫩江流域青铜时期人群生业模式和文化改变的原因是否与外来人群进入有关,我们将嫩江流域的古代人群分成新石器时代和青铜铁器时代两组人群进行随后的人群遗传分析。
表2 嫩江流域古代人群线粒体全基因组检测结果
Tab.2
| 样本编号 | 考古遗址 | 线粒体测序深度 | 线粒体污染率 | 线粒体单倍群 |
|---|---|---|---|---|
| HT-M45 | 后套木嘎遗址 | 853.3641 | 0.008 | D4h1+12396C |
| HT-M89 | 后套木嘎遗址 | 68.0831 | 0.016 | A+152C |
| HT-M80 | 后套木嘎遗址 | 55.223 | 0.012 | D4e5a |
| HT-M91 | 后套木嘎遗址 | 28.4945 | 0.032 | Y1a |
| HT-M54A | 后套木嘎遗址 | 21.8996 | 0.058 | B4c1a2 |
| HT-M94 | 后套木嘎遗址 | 4.7071 | 0.037 | D4c1b |
| HQHM2 | 洪河村遗址 | 52.0482 | 0.008 | D4c1b |
| HQHM3 | 洪河村遗址 | 17.6003 | 0.009 | D4j |
| HQHM4 | 洪河村遗址 | 59.4463 | 0.022 | D4b2a |
| HQHM5 | 洪河村遗址 | 20.8831 | 0.016 | D4 |
| ZLNR-2 | 扎赉诺尔遗址 | 16.6342 | 0.014 | C5 |
| WQM4 | 五七农场遗址 | 16.9741 | 0.015 | C4a1a |
| HT-M74A | 后套木嘎遗址 | 72.9727 | 0.019 | C4a2a1 |
| HT-M25B | 后套木嘎遗址 | 46.2914 | 0.034 | D4c2b |
| HT-M69B | 后套木嘎遗址 | 44.3536 | 0.092 | G2a1 |
| HT-M74B | 后套木嘎遗址 | 30.9066 | 0.017 | G2a1 |
| HT-M69A | 后套木嘎遗址 | 23.8907 | 0.046 | B4c1a2 |
| HT-M69C | 后套木嘎遗址 | 5.6121 | 0.042 | F1b1e |
| DGDM104 | 大古堆遗址 | 59.44631 | 0.026 | M8* |
| DGDM105 | 大古堆遗址 | 20.883 | 0.01 | D4m |
| ZLNR-1 | 扎赉诺尔遗址 | 5.7467 | 0.012 | N9a9 |
| MGS-M6 | 内蒙古蘑菇山 | 19.3666 | 0.009 | C5a1 |
| MGS-M7L | 内蒙古蘑菇山 | 22.2542 | 0.006 | Z3a1 |
| MGS-M7R | 内蒙古蘑菇山 | 33.9752 | 0.015 | C4a1a4a |
早期人群(n=12)主要携带有单倍群A、B4、C4、C5、D4、Y,包括12个亚单倍群,展现了较高的人群内部遗传多样性。单倍群D4是早期人群中最为主要的单倍群(n=7),频率高达58.33%,其在现代的北亚和远东地区以及古代的北亚人群中有着非常高的频率(图2)[13],在现代的中国北方人群中也有着较高的频率(19.47%)[15]。嫩江流域早期人群的整体单倍群构成与远东地区新石器时代的古代人群和蒙古新石器时代古代人群较为相似,都有高频的单倍群D和C(图2)。与早期人群相比,晚期人群(n=12)展现了更为复杂的单倍群多样性,包括出现在西辽河古代人群以及黄河流域古代人群的单倍群N9a、Z、M、F,发现于贝加尔湖和西辽河古代人群中G谱系。嫩江流域晚期人群单倍群构成与西辽河地区新石器时代人群和黄河流域新石器时代古代人群以及现代当地少数民族更为相似(图2)。
图2
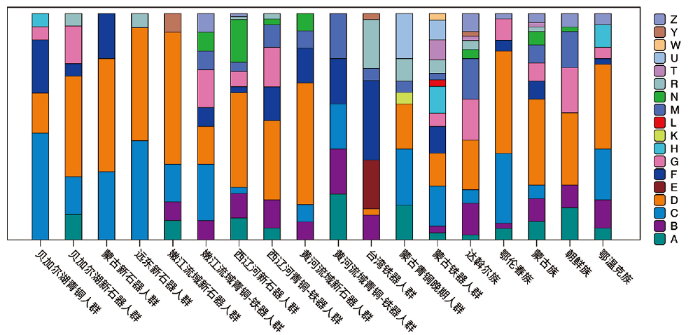
图2
嫩江流域古代人群与其他古代及现代人群线粒体单倍群频率示意图
横坐标为各个比较人群,纵坐标为单倍型频率,不同的颜色代表不同的线粒体单倍群
Fig.2
Relationship between ancient NenJiang River populations and other populations based on haplogroup frequencies
The abscissa is the comparison populations, and the ordinate is the frequency of haplogroups. Different colors represent different haplogroups of mitochondria DNA
为了进一步揭示嫩江流域古代人群与周边古代人群的关系,我们基于线粒体单倍群频率构建了主成分分析(PCA)。在PCA图中,从第一主成分和第二主成分(2个主成分共占比例为33.7%)可以看出,所分析的古代人群的分布大体上与其地理分布相吻合,来自相近地区的古代人群在PCA中具有相似的遗传分布。其中,嫩江流域早期人群与远东古代人群遗传距离最近(他们主要共享的单倍群是D4、C5、A+152C等),而嫩江流域晚期人群在第一主成分上与早期人群分开,反而与黄河流域新石器时代人群以及西辽河地区古代人群聚在一起(主要受单倍群N9a、Z、D5和G的影响)(图3)。主成分分析表明,嫩江流域早晚期人群母系遗传结构发生了动态变化。
图3
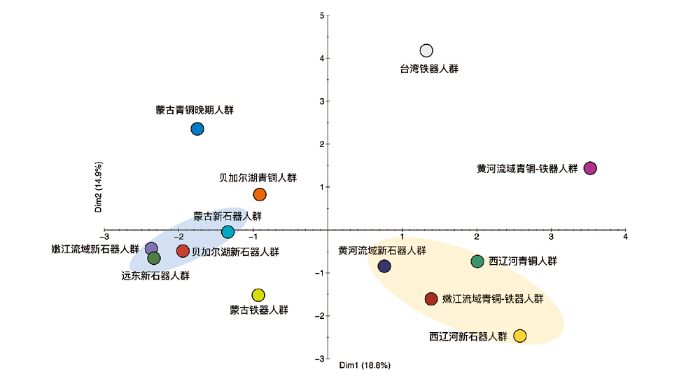
图3
嫩江流域古代人群与相关古代人群主成分分析
嫩江流域早期人群所处的遗传聚类用浅蓝色表示,嫩江流域晚期人群所处的遗传聚类用浅黄色表示
Fig.3
PCA plot based on haplogroup frequencies of Nenjing River populations and other ancient populations calculated using
The genetic cluster of the early Nenjiang River population is shown in light blue and the genetic cluster of the later Nenjiang River population is shown in light yellow
为了详细探讨这种遗传结构变化的程度,我们对早晚期人群的更下游的单倍群进行了分析。我们发现,嫩江流域早晚期人群共享单倍群C4a1a、C5以及B4c1a2。C4a1a主要分布在古代的蒙古人中,从新石器至青铜铁器均有发现[12],其次,在青铜贝加尔湖人群以及中国北方黄河流域仰韶人群也有存在[9,13];单倍群C5则特异且高频的分布在古代俄罗斯远东人群中[13];而单倍群B4C1a2只在嫩江流域和西辽河地区古代人群中被发现[9],暗示这些单倍群可能起源于东北亚地区。结合已发表的常染色体基因组结果,我们推测这些单倍群在早晚期人群中的共存,可能是嫩江流域人群长期的遗传连续性和稳定性导致的,并且这种遗传连续性可能一直持续到当地的现代人群,比如辽宁、吉林、黑龙江、内蒙古的现代人群中也有低频的C4a1a、C5以及B4c1a2 [15]。为了进一步追踪这种遗传连续性,我们针对数据质量较好的单倍群B4c1a2执行了中介网络分析(图4A)。中介网络图显示,嫩江流域早期的后套木嘎古代样本(HT-M54A,7500BP)与晚期的后套木嘎古代样本(HT-M69A,2500-2000BP)在同一分支上,并且早期的样本处于系统发育树的根部,表明后套木嘎早晚期人群在母系遗传上是连续的。此外,后套木嘎早期样本与西辽河地区青铜时期的一个样本(91KLM2,2800-2300BP)以及现代雅库特的一个个体处于另一个分支上(图4A)。值得注意的是,尽管91KLM2这个样本所处的地理位置在西辽河地区,其全基因数据却显示与嫩江流域的人群聚在一起,表明这个样本所代表的个体起源于嫩江流域[9]。因此,中介网络分析支持嫩江流域古代人群自7500至2000年左右具有母系遗传连续性。
图4
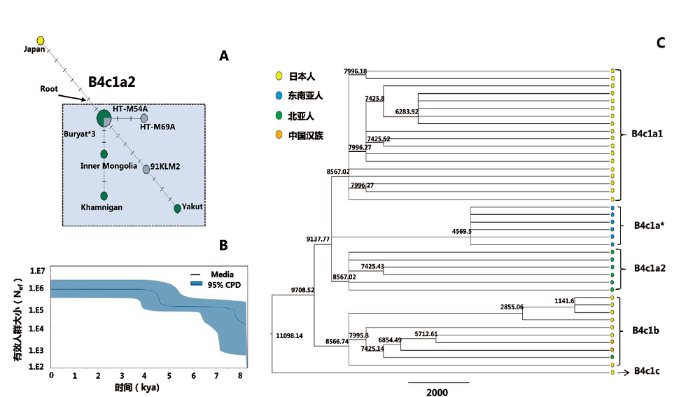
图4
基于单倍群B4c1a序列的统计学分析
A)单倍群B4c1a2的中介网络图/The network of haplogroup B4cla2;B)单倍群B4c1的贝叶斯Skyline曲线 The Bayesian Skyline plots of Haplogroup B4c1 showing the maternal effective population size history;C)单倍群B4c1序列的贝叶斯树,分支上的数字代表分支的分歧时间,其中B4c1a2的共祖时间是大约距今8567年
Fig.4
Statistical analyses based on the sequences of Haplogroup B4c1a
The Bayesian tree for sequences from Haplogroup B4c1. The long of line is the years before present, the number is the age for the divergence age for the clade and the coalescence time of B4c1a2 was 8560 years ago, the individuals are colored according to different origin
为了考察嫩江流域的人群历史动态,我们应用DnaSP软件基于线粒体DNA序列对新石器(n=12)和青铜铁器(n=12)两个时期人群分别执行了中性检验,结果Tajima’D均为负值(新石器时期;-0.8488(P>0.1);青铜铁器时期:-1.0438(P>0.1)),但检验结果均不显著,说明从新石器时期开始一直到青铜铁器时期,嫩江流域人群保持相对的稳定性,并没有发生明显的人群扩张或者衰减事件。为了进一步追踪嫩江流域史前人群历史动态,我们基于单倍群B4c1a的线粒体全基因组数据重构了嫩江流域史前人群历史。EBSP曲线表明线粒体单倍群B4c1a分别在距今9000-7500年和距今5000-4000左右有一个人口增长期(图4B)。第一次扩张时间与伴随着全新世大暖期开始北方以旱作农业为特点的新石器文化的扩张时间想吻合[18]。第二次发生扩张的时间与龙山时期粟作农业的扩张时间有很好的应对[19]。共祖模拟分析显示B4c1a2的共祖时间大约是在8500年左右(图4C),表明距今8500 年前,B4c1a2就已经存在,并于7500年前扩散到了嫩江流域,与单倍群B4c1a第一次的扩张时间相一致。
尽管嫩江流域早晚期人群在全基因组水平和线粒体水平上均展现了较强的遗传连续性,但是,线粒体数据同时也展现了非常明显的遗传差异。PCA图显示,嫩江流域晚期人群与中原黄河流域新石器时代人群以及西辽河古代人群有着较近的遗传距离(图3),这可能受单倍群Z3和N9a的影响。不论是单倍群Z3a1还是其上游Z3目前在所获得的整个东北亚地区古代人群中只在西辽河地区夏家店下层存在。Z3a1在现代人群中的频率也较低,主要分布在中国北方人群。因此嫩江流域晚期人群中出现的Z3a1单倍群不太可能来源于东北亚地区,而更可能是来源于西辽河地区。而晚期人群中出现的单倍群N9a(ZLNR-1)在辽西地区新石器时代的朝阳半拉山人群(BLS41)和青铜时期夏家店下层文化的二道井子人群(EDM176)中均有发现,同时N9a在中原瓦店龙山文化人群中(WT1H16个体)以及黄河上游铁器时期大槽子人群中也有低频存在[9](表2)。此外,赵欣等人对西辽河古代个体的线粒体DNA高可变区序列分析显示,N9a在西辽河红山文化的牛河梁遗址的频率高达35.71%,在小河沿文化的哈啦海沟人群中占8.33%[11],而在嫩江流域的早期古代人群以及蒙古、贝加尔湖和俄罗斯远东的古代人群中均未发现N9a [20,21,22]。单倍群N9a在古代人群中的分布模式表明N9a很可能起源于西辽河地区。西辽河来源的单倍群Z3a1和N9a在嫩江流域晚期人群中的存在表明,西辽河古代人群曾向嫩江流域扩散,并留下母系遗传印迹。为了进一步追踪这种基因流向和扩散历史,我们基于线粒体基因组数据对于数据质量相对较好的单倍群N9a做了共祖模拟分析、BESP分析和中介网络分析。EBSP曲线表明线粒体单倍群N9a的有效人群自距今6000-4000BP有个扩张期(图5A,5B)。古植物学研究表明,距今6000-4000年前正是粟黍农业在中国北方广泛扩张,推动新石器晚期文化的繁盛和人口显著增长的时期[23]。此外,考古学研究表明,距今6000年前正是辽西地区红山文化兴起的时间,而4000年左右则是中原龙山文化晚期中原农业人群向辽西地区扩张的时期[8,9],单倍群N9a人口数量扩张期与辽西地区本土新石器中晚期文化发展是相一致的。同时,我们构建了N9a的中介网络图(西辽河古代个体BLS41数据质量较差,在此分析时排除)试图探讨各个地区N9a个体之间的关系。中介网络图显示(图5C),单倍群N9a在现代人群中广泛分布在中国、日本、北亚以及东南亚。其中日本的现代样本大部分与黄河流域龙山时期的古代样本(WT1H16)聚在一个分支,并且古代样本在根部,暗示这一分支可能起源于中国。而西辽河青铜时代样本EDM176与嫩江晚期ZLNR-1样本并不在一个分支,并且他们所处分支个体数量有限,无法判断他们的具体起源地。但是,结合单倍群N9a在古今人群中的存在和分布频率以及N9a扩张时间与西辽河地区考古学文化的对应关系,我们可以推测N9a在嫩江流域晚期人群中的存在与辽西地区的古代人群向嫩江流域迁徙扩张有关。
图5
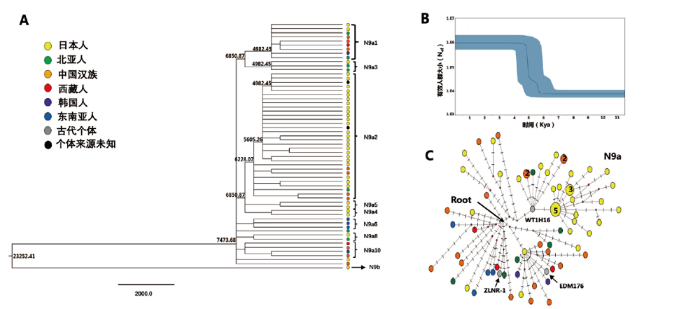
图5
基于单倍群N9a序列的统计学分析
(A) 单倍群N9a序列的贝叶斯树,分支上的数字代表分支的分歧时间, 不同地理来源的个体应用不同的颜色表示,每个个体的Genbank 号见附
Fig.5
Statistical analyses based on the sequences of Haplogroup N9a
(A)The Bayesian tree for sequences from Haplogroup N9a. The long of line is the years before present, the number is the age for the divergence age for the clade, the individuals are colored according to different origin, the Genbank number for every individual was shown in Supplementary Fig S3. (B) The Bayesian Skyline plots of Haplogroup N9a showing the maternal effective population size history. (C)The Network of Haplogroup N9a. Haplotypes where the number of Shared individuals is greater than one are numbered
4 讨论
考古调查表明,松嫩平原新石器早期的制陶工艺以及纹饰风格受到俄罗斯外贝加尔地区卡棱加河口文化的影响,与贝加尔湖地区存在高度的相似性[5]。正如母系遗传数据所示,早期的松嫩人群与蒙古新石器时代和北亚新石器时代的渔猎人群聚在一起,与俄罗斯远东人群展现了非常相似的遗传特点。这一结果表明新石器早期,松嫩平原渔猎人群应该与其同纬度以及北部的渔猎人群有着相似的遗传基质和文化内涵。青铜时期以后,人群母系遗传成分上与早期人群具有一定程度的遗传连续性同时,也发生了一定的变化。嫩江地区晚期古代人群(4000-2000)与西辽河地区青铜铁器时期人群(4000-2000)以及中原新石器时代人群的遗传结构更为相似。结合单倍群Z3a1和N9a谱系的起源以及N9a的扩张时间,我们认为西辽河地区古代人群在距今2000年之前曾向松嫩平原迁徙,并留下了母系遗传印迹。值得指出的是,Ning等人对松嫩平原古代人群的5个个体(7500-2000)全基因组研究显示出该地区人群有较强的遗传连续性,早晚期人群均与北亚现代人有着较近的遗传关系,并没有发现西辽河遗传成分的进入,只有晚期的一个个体显示出部分蒙古成分的融入[9]。线粒体DNA基因组不但追踪到了这种遗传连续性,同时还发现了外来人群对当地人群母系遗传结构的影响,这可能归因于我们采用了更大的样本量以及线粒体DNA可以呈现更高的人群内部差异性。
此外,PCA图显示,西辽河青铜铁器时代人群较新石器时代人群与中原新石器时代人群关系更为密切,揭示西辽河青铜铁器时代人群与红山文化时期的人群的遗传差异主要来自于中原农业人群向西辽河地区的扩张。这与西辽河地区夏家店下层农业文化、陶器风格等具有明显的中原文化影响是相一致的[24]。而嫩江流域晚期人群在PCA上介于西辽河青铜铁器时代人群与中原新石器时代人群之间,与二者均具有较近的遗传距离可能表明,西辽河古代人群向松嫩平原的迁徙可能发生在中原农业人群向西辽河地区扩张并与当地人群发生融合之后。考古调查表明,夏家店下层文化时期(4000-3500BP),西辽河地区考古学文化与中原考古学文化有着密切的联系[23], 而这一时期也刚好是嫩江流域考古文化发生重要转变的时间,因此我们推测西辽河地区古代人群向松嫩地区的迁徙扩张应该是在4000-3500年间。
古环境学研究表明,距今5000-3000年,东北地区气候开始持续变冷[25]。而这一时期,正是中原发达农业文化强势向西辽河地区扩张的时期,中原人群的进入给西辽河地区带来了先进农业技术的同时,也伴随着因人口的快速增长带来的资源压力[9]。这种生存压力迫使西辽河古代人群向嫩江流域迁徙并给当地带去了新的技术与语言文化。这与语言学上认为的,盛行于嫩江流域的泛欧亚语言来源于西辽河地区是相一致的(一般认为语言的传播是伴随着人群迁徙的[26])。而松嫩平原古代人群可能为了适应因气候恶劣而造成的渔猎采集资源不足,对生业模式做出了调整,让适量的农业和畜牧业加入作为辅助的生业模式。这一时期文化的复杂度以及各种生业模式并存的现象可能正是由于当地的人群为了适应气候变化所做出的改变。考虑到嫩江流域古代人群全基因组数据和线粒体基因组数据均展现了较强的遗传连续性,辽西地区古代人群向嫩江流域的人群迁徙规模可能不大。
参考文献
Bioarchaeological perspective on the expansion of Transeurasian languages in Neolithic Amur River basin
[J].
Ancient genomes from northern China suggest links between subsistence changes and human migration
[J].DOI:10.1038/s41467-020-16557-2 URL [本文引用: 12]
The Genomic Formation of Human Populations in East Asia
[J].
The first horse herders and the impact of early Bronze Age steppe expansions into Asia
[J].DOI:10.1126/science.360.6396.1391 URL PMID:29954963 [本文引用: 4]
Investigating Holocene human population history in North Asia using ancient mitogenomes
[J].
River Valleys Shaped the Maternal Genetic Landscape of Han Chinese
[J].
DOI:10.1093/molbev/msz072
URL
PMID:31112995
[本文引用: 3]

A general south-north genetic divergence has been observed among Han Chinese in previous studies. However, these studies, especially those on mitochondrial DNA (mtDNA), are based either on partial mtDNA sequences or on limited samples. Given that Han Chinese comprise the world's largest population and reside around the whole China, whether the north-south divergence can be observed after all regional populations are considered remains unknown. Moreover, factors involved in shaping the genetic landscape of Han Chinese need further investigation. In this study, we dissected the matrilineal landscape of Han Chinese by studying 4,004 mtDNA haplogroup-defining variants in 21,668 Han samples from virtually all provinces in China. Our results confirmed the genetic divergence between southern and northern Han populations. However, we found a significant genetic divergence among populations from the three main river systems, that is, the Yangtze, the Yellow, and the Zhujiang (Pearl) rivers, which largely attributed to the prevalent distribution of haplogroups D4, B4, and M7 in these river valleys. Further analyses based on 4,986 mitogenomes, including 218 newly generated sequences, indicated that this divergence was already established during the early Holocene and may have resulted from population expansion facilitated by ancient agricultures along these rivers. These results imply that the maternal gene pools of the contemporary Han populations have retained the genetic imprint of early Neolithic farmers from different river basins, or that river valleys represented relative migration barriers that facilitated genetic differentiation, thus highlighting the importance of the three ancient agricultures in shaping the genetic landscape of the Han Chinese.
The genome sequence of a 45,000-year-old modern human from western Siberia
[J].
DOI:10.1038/nature13810
URL
PMID:25341783
We present the high-quality genome sequence of a approximately 45,000-year-old modern human male from Siberia. This individual derives from a population that lived before-or simultaneously with-the separation of the populations in western and eastern Eurasia and carries a similar amount of Neanderthal ancestry as present-day Eurasians. However, the genomic segments of Neanderthal ancestry are substantially longer than those observed in present-day individuals, indicating that Neanderthal gene flow into the ancestors of this individual occurred 7,000-13,000 years before he lived. We estimate an autosomal mutation rate of 0.4 x 10(-9) to 0.6 x 10(-9) per site per year, a Y chromosomal mutation rate of 0.7 x 10(-9) to 0.9 x 10(-9) per site per year based on the additional substitutions that have occurred in present-day non-Africans compared to this genome, and a mitochondrial mutation rate of 1.8 x 10(-8) to 3.2 x 10(-8) per site per year based on the age of the bone.
Paleolithic to Bronze Age Siberians Reveal Connections with First Americans and across Eurasia
[J].DOI:10.1016/j.cell.2020.03.025 URL PMID:32243785 [本文引用: 1]
A dynamic 6,000-year genetic history of Eurasia’s Eastern Steppe
[J].
The population history of northeastern Siberia since the Pleistocene
[J].
DOI:10.1038/s41586-019-1279-z
URL
PMID:31168093
[本文引用: 1]
Northeastern Siberia has been inhabited by humans for more than 40,000 years but its deep population history remains poorly understood. Here we investigate the late Pleistocene population history of northeastern Siberia through analyses of 34 newly recovered ancient genomes that date to between 31,000 and 600 years ago. We document complex population dynamics during this period, including at least three major migration events: an initial peopling by a previously unknown Palaeolithic population of 'Ancient North Siberians' who are distantly related to early West Eurasian hunter-gatherers; the arrival of East Asian-related peoples, which gave rise to 'Ancient Palaeo-Siberians' who are closely related to contemporary communities from far-northeastern Siberia (such as the Koryaks), as well as Native Americans; and a Holocene migration of other East Asian-related peoples, who we name 'Neo-Siberians', and from whom many contemporary Siberians are descended. Each of these population expansions largely replaced the earlier inhabitants, and ultimately generated the mosaic genetic make-up of contemporary peoples who inhabit a vast area across northern Eurasia and the Americas.
Evolution of human-environmental interactions in China from the Late Paleolithic to the Bronze Age
[J].
Millet agriculture dispersed from Northeast China to the Russian Far East: Integrating archaeology, genetics, and linguistics
[J].


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}